By G Miller, R Cummings, A Hester
Summary
Many of the few remaining patches of natural woodland in the Cairngorms are failing to regenerate because tree and shrub saplings are repeatedly checked or killed by browsing red deer. Liability to browsing is governed by a complex of interacting factors including a sapling’s physical and chemical properties, season of year, availability of alternative foods, soil conditions, nature of surrounding vegetation and local red deer density. Amongst common native tree and shrub species in the Cairngorms, deer prefer to eat rowan but generally take juniper only when alternative foods are scarce. Young Scots pine saplings are the most easily killed by browsing whereas birch and rowan can survive repeated damage. Many saplings, including pines, are maintained in a suppressed state by repeated browsing. These plants constitute an often long-lived ‘sapling bank’ from which rapid growth can occur once browsing pressure has been relieved.
On unburnt ground in the Cairngorms, conditions for tree regeneration are currently most favourable at altitudes greater than 550 m because here there are (a) fewer deer and (b) more gaps for seedling establishment than is the case on lower ground. Reducing red deer density to fewer than about 5 animals per km2 increases the possibility of woodland regeneration everywhere. However, this in itself may be insufficient to produce good seedling establishment in dense ericoid or graminoid vegetation with its associated deep mor humus layer. Ground preparation by fire or by mechanical disturbance may be necessary in such circumstances if rapid and extensive regeneration is required.
Introduction
Natural and semi-natural scrub and woodland presently occupy less than 10% of the 5 000 km2 Cairngorms Partnership Area, considerably less than the supposed maximum cover of around 80% (Cairngorms Working Party, 1992). This deficiency has resulted from nearly four centuries of felling, burning and grazing which first destroyed most of the original forest and then prevented its regeneration (Carlisle, 1977). The ecological consequences have involved significant changes to microclimate, soils, flora and fauna. Now there is increasing interest in the possibility of restoring natural woodland cover (e.g. Beaumont etal, 1995). Indeed, the potential extent and nature of reconstructed native woodland in the region has recently been modelled by Macmillan et al. (1997) who concluded that pine and oak-birch might be the most extensive types.
At present, saplings of Scots pine (Pinus sylvestris), birch (Betula pendula and B. pubescens), rowan (Sorbus aucuparia), alder (Alnus glutinosa), willows (Salix spp.) and juniper (Juniperus communis) are scattered throughout the Cairngorms. Pine and birch saplings can be locally plentiful, given a seed source and suitable ground conditions, but the other species are generally scarce. Large blocks of ground below the 600-650 m tree-line are largely bereft of saplings. Where they do occur, repeated browsing usually restricts their height to no more than a few centimetres.
Wild and domesticated herbivores continuously check tree regeneration over most unfenced land in the Cairngorms. The chief agents are red deer (Cervus elaphus), though sheep, cattle, roe deer (Capreolus capreolus), mountain hares (Lepus timidus), rabbits (Oryctolagus cuniculus), voles (Microtus agrestis) and various invertebrate species also contribute. Herbivore predation of native tree and shrub saplings has been little researched and most of the work that has been done concerns red deer. Their great mobility, wide dispersion and liking for browse make them most effective destroyers of regenerating trees.
Scrub and woodland regeneration in the Scottish uplands as a whole has been discussed in detail by Hester and Miller (1995) and by Hester (1995). Gill (1992a and b) has considered the impact of mammals in northern forests generally. Our review is less comprehensive and focuses on the impact of red deer on tree establishment, survival and growth in the Cairngorms. It addresses four key questions:
- What determines a sapling’s liability to browsing?
- How tolerant of browsing are saplings of different species?
- Would a simple reduction in red deer numbers allow woodland regeneration?
- If so, what other factors control the nature and longevity of succession to woodland?
Liability to Browsing
Factors controlling diet selection by deer are diverse and complex. The attractiveness of any sapling is determined primarily by its physical and chemical characteristics. Both palatability and digestibility vary amongst species, depending on nutrient content and the presence of secondary compounds (Bryant et al, 1991; Tahvanainen et al, 1991). Tissues with an elevated nitrogen content seem especially attractive (Rusterholz and Turner, 1978; Niemela and Danell, 1988) and the application of nitrogenous fertiliser to Scots pine has been shown to encourage browsing damage (Laine and Mannerkoski, 1980). In general, saplings rooted in any relatively fertile soil are vulnerable, not only because of their enhanced nutrient content, but also because they are located amongst attractive herbage (Danell, 1989; Danell et al., 1991). Hence tree establishment and survival tends to be especially poor amongst the short grassy vegetation characteristic of the relatively fertile soils on the floors of some glens in the Cairngorms.
Some woody species produce what are thought to be anti-herbivore chemicals. A prime example is juniper which contains oxygenated monoterpenes. These act by rendering the plant tissues distasteful and by interfering with an animal’s digestion (Schwartz et al., 1980a and b). Scots pine shoots also contain monoterpenes (Forrest, 1980), whereas aspen (Populus tremula), rowan and willows, all heavily browsed in the Cairngorms, seem much less well defended (Tahvanainen et al., 1991).
The attractiveness of different species also varies seasonally. On wintering ground at Glen Feshie, red deer seldom took leafless birch saplings during winter but they actively selected them at bud burst in May-June and continued browsing until leaf fall in October (Miller and Cummins, 1998). By contrast, evergreen but monoterpene-rich juniper saplings were generally avoided during summer and taken mainly in winter. In fact the main period of juniper browsing at that site was in March-April, when alternative foods had become scarce.

Red deer will browse most tree and shrub species at one time or another, but there is a distinct hierarchy of preference (Mitchell et al., 1977; Gill, 1992a; Hester and Miller, 1995). Precise species rankings can vary according to season and to local site conditions. However, field observations (Cummins and Miller, 1982; Mitchell et al., 1982) and experimental plantings (Miller et al., 1982) suggest that the general sequence amongst common trees in the Cairngorms area is rowan > willow > pine and birch > juniper. Aspen too is highly preferred but is relatively rare in the area.
Liability to browsing is also a function of site characteristics. These include the species composition, density and height of the vegetation amongst which saplings grow. For instance, birch sapling survival in the Cairngorms is much superior amongst heather (Calluna vulgaris) than it is amongst relatively palatable grassy vegetation (Kinnaird, 1974; Stewart, 1996). This is also true of pine saplings (Palmer et al., 1998). Vegetation height and density affect the conspicuousness of saplings. Those projecting above the general level of the sward usually have little chance of escaping damage where large numbers of deer are active (Miller et al., 1982). Even the density, height and species composition of a sapling population itself influences the severity and seasonality of browsing. Damage to Scots pine tends to be greatest where sapling density is sparse and most severe on individuals 1-2 m tall (Cummins and Miller, 1982). In mixed stands with deciduous species predominant, pines are generally browsed mostly in winter; but where deciduous species are scarce, they are taken mainly in summer (Cummins and Miller, 1982).
Herbivore density is the prime site factor governing liability to browsing. In the 1995 census, 32 010 red deer were counted on the 240 000 ha of the Cairngorms/West Grampians area, giving an overall density of 13 per km2 (Youngson and Stewart, 1996). Such a figure is of little value in evaluating deer impact because their local abundance varies widely – spatially, diurnally and seasonally. Accurate information on local deer abundance is scant, although rough estimates can be made from dung deposition (Mitchell and McCowan, 1980; Mayle and Staines, 1998). For instance, dung counts in 1969- 70 indicated densities of 0.2-1.7 deer per ha on different parts of wintering ground at Glen Feshie (Miller and Cummins, 1998). These variations in deer density were linearly related to birch sapling predation, with 1 deer per ha being roughly equivalent to 2% of saplings browsed weekly. However this relationship would alter in summer, when birch is in leaf and is actively selected.
Various attempts have been made to estimate the density of red deer which might allow succession to woodland in the Scottish uplands. These are partly conjectural but do show a degree of consistency in ranging from 2 to 10 deer per km2 (Table 1). Regeneration, however, is mediated by so many diverse and interacting factors that such figures need to be interpreted with care. They also need to reflect real situations. For example, the estimate of 10 deer per km2 to allow tree establishment assumes a starting density of at least 4 000 saplings per ha (Table 1), although recorded sapling densities seldom exceed 1 000 plants per ha in the Cairngorms (Miller and Cummins, 1982; Beaumont et al., 1995; French et al., 1997b). As discussed, much also depends on the species and distribution of the saplings, the availability of other browse, the vegetation type, soil fertility, and herbivore distribution. Even so, reducing red deer density to around 5 or fewer per km2 resulted in a 25% increase in sapling density at Abernethy (Beaumont et al., 1995) and allowed checked saplings to grow at Inshriach and at Creag Meagaidh (Staines et al., 1995).
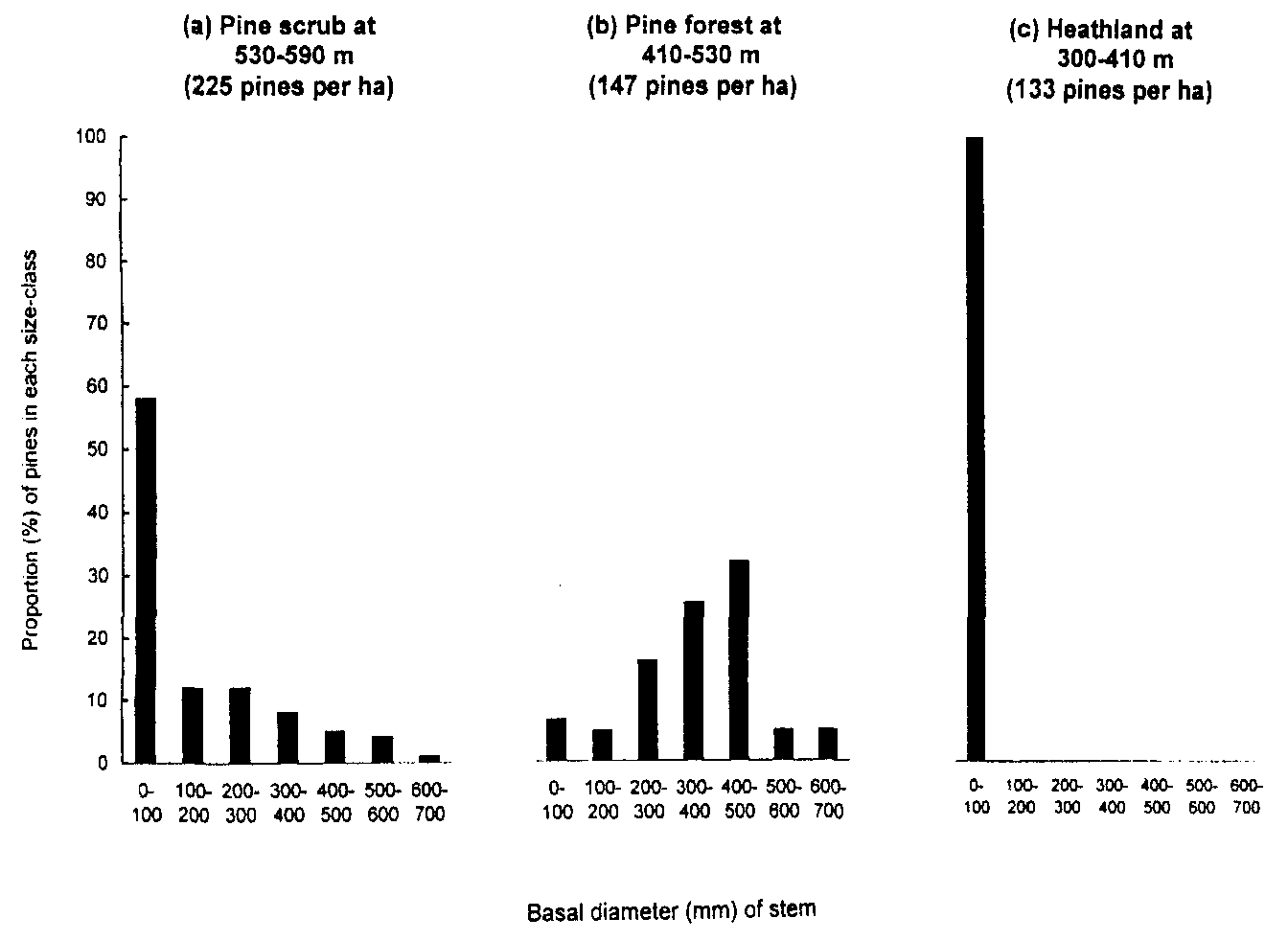
Stocks of red deer on low ground are usually much greater than the 10 animals per km2 proposed by Mitchell et al. (Table 1) and locally they often exceed 100 animals per km2 in winter (Youngson and Stewart, 1996). However, deer densities generally decline with increasing altitude, especially in winter, and there is a parallel decline in the liability of saplings to browsing (Miller et al., 1982). Hence the best prospects for sapling survival in the Cairngorms may Currently be around the assumed tree line of 600-650 m. Indeed the size structure of pine populations on Creag Fhiaclach in the early 1970s suggests that whilst saplings at the tree line could develop to maturity, those on lower ground were kept in check by repeated browsing (Figure 1).
Tolerance of Browsing
Plant response to browsing is dependent on the frequency, severity and season of damage in relation to the size and age of the individual. Mature trees are generally least vulnerable and large individuals can withstand even severe damage to the lower branches (Bryant et al., 1985). Hence, well-trimmed old pines, birches, willows and junipers are a common sight, especially in the western Cairngorms. Damage to young trees up to 3 m tall may result in reduced and distorted growth, desiccation, or infection by pathogens. Saplings and seedlings less than 30 cm tall are easily stunted or even killed by browsing. In general, clipping studies (reviewed by Mitchell et al., 1977; Canham et al., 1994; Ouellet et al., 1994; Edenius et al., 1994) show that sapling mortality increases with the frequency of damage and with the amount of tissue removed. Probability of survival is least when damage occurs during the growing season (Canham et al, 1994). Survival is also a function of soil fertility, with good nutrition enhancing a sapling’s chances of a successful recovery after damage (Danell et al., 1991; Edenius et al., 1994). However, in much of the Cairngorms soils are inherently infertile (Heslop, 1981), browsing is recurrent and, consequently, most saplings remain stunted or are killed.
Tolerance of browsing clearly varies with plant species. Pines and other conifers tend to be particularly sensitive to severe damage because much of their carbon and nutrient reserve is located in the needles (Chapin, 1980). Experimental clipping has shown that two-year-old pine saplings inevitably died if cut back into the previous season’s wood, whereas most birch and all rowan survived (Miller et al, 1982). Edenuis et al., (1994) also caused heavy mortality amongst young Scots pine by removing 90% of the needle biomass. Their vulnerability to defoliation probably contributes to the notable scarcity of pines on the fertile soils and short grassy vegetation of glen floors, as in Glen Feshie (Miller and Cummins, 1998). Nevertheless pine saplings commonly survive on ground frequented by red deer in the Cairngorms, especially if growing on podzolic soils amongst tall heather or other protective vegetation. The other native conifer of the Cairngorms, juniper, has not been clipped experimentally but field evidence (Fitter and Jennings, 1975) suggests that the saplings can in fact withstand repeated and severe browsing. Their comparative rarity, particularly in the eastern Cairngorms, is more likely due to sensitivity to fire (Hobbs et al., 1984).
Deciduous species generally tend to be much less sensitive to browsing. Thus annual mortality rates of 7% and 5% have been recorded respectively for browsed birch and rowan saplings in Glen Feshie as compared with 20% for pines (J.W. Kinnaird, unpub). Indeed birch is so resilient that repeatedly browsed plants a few centimetres tall can be up to 31 years old when growing amongst heather (Kinnaird, 1974). This could explain why birch and rowan are so over-represented in the sapling flora of Glen Feshie compared with their relative scarcity amongst the mature trees in the pine woodland there (Miller and Cummins, 1998).
Sources of Regeneration
If red deer density is sufficiently low, forest succession in the Cairngorms can come from either or both of two sources.
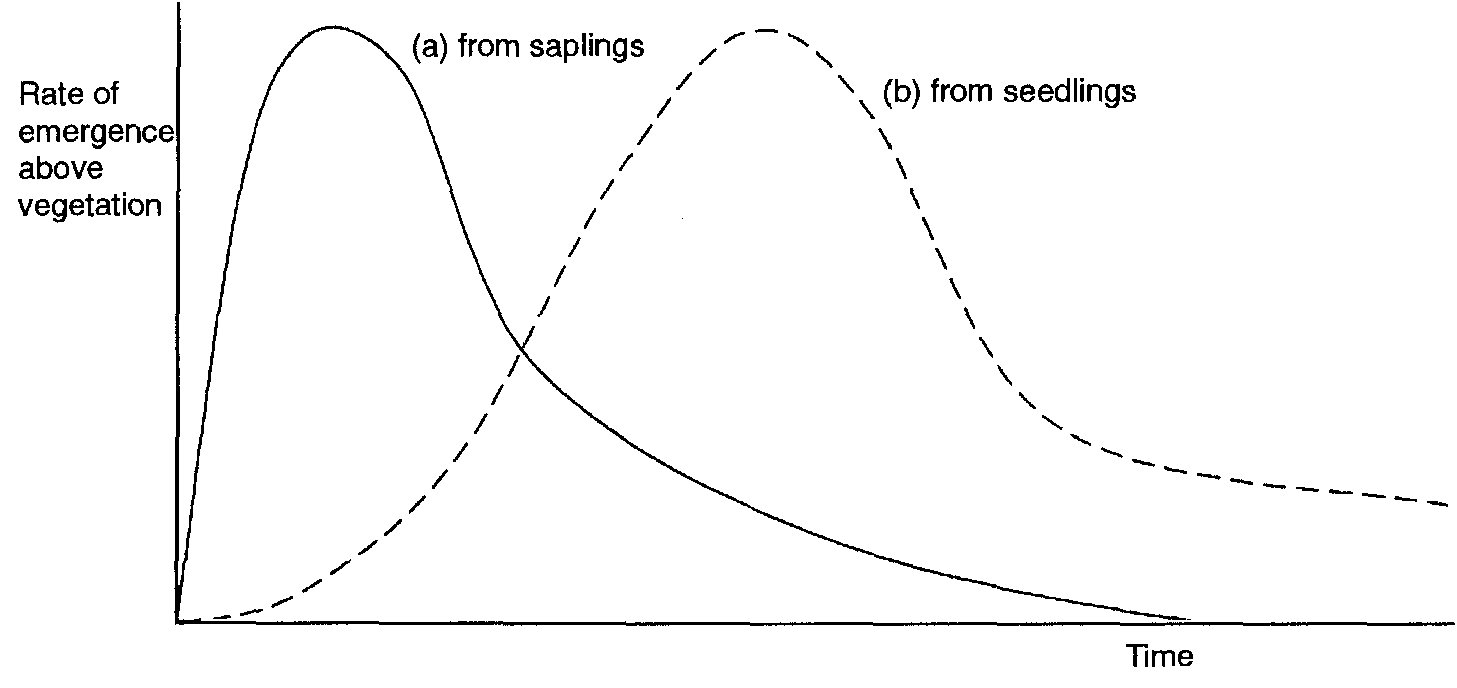
These are (a) the rapid growth of long-established but suppressed saplings, and (b) the slow establishment of new plants from seeds. Where both sources are available, the pattern of the succession may be bimodal (Figure 2). The balance between the two types of regeneration at any particular site will depend on its environmental history.
(a) Regeneration by growth of long-established saplings
Many potential woodland sites in the Cairngorms already have long-established populations of saplings held in check by persistent browsing. The few examples that have been quantified show densities ranging up to 900 saplings per ha (e.g. Beaumont et al., 1995). In effect, this reservoir of suppressed saplings is analogous to the so-called ‘seedling bank’ of shade-tolerant species characteristic of tropical forests (Whitmore, 1990). Just as a tropical forest seedling bank responds to increased light, so the suppressed saplings in the Cairngorms will respond to decreased browsing pressure from red deer.
A substantial reduction in deer numbers should result in the immediate growth of any resident suppressed saplings. Just such a response seems to have been observed at Abernethy where recorded sapling numbers increased by 20% three years after deer reduction (Beaumont et al., 1995), at Inshriach where they increased three-fold in four years, and at Creag Meagaidh where they increased about four-fold in three years (Staines et al., 1995). Other examples of growth by formerly checked saplings can be seen in small fenced plots at Glen Feshie, Glen Derry and elsewhere in the Cairngorms.
A well-established sapling bank will be more influential than seed fall in determining the initial character of the new woodland (Figure 2). Indeed its initial composition may be quite different from that of surrounding mature woodland.
For example, Miller and Cummins (1998) found that birch comprised 84% of the sapling bank within their sample areas at Glen Feshie, even though the adjacent woods were dominated by planted and self-sown Scots pine. Pines in fact accounted for less than 1% of the sapling bank, presumably because they are more easily killed by browsing than are birches. Hence, in some parts of the Cairngorms, a reduction in deer browsing pressure may result initially in woodland with an unexpectedly high proportion of broadleaved trees.
(b) Regeneration by establishment of new seedlings
Recent events in the northern Cairngorms illustrate the early stages of forest succession by seedling establishment (French et al., 1997b). Young pines have colonised the northern corries of Cairn Gorm along an altitudinal gradient from the edge of mature forest at 450-500 m up to around 850 m. About 90% of the saplings were aged less than 25 years in 1985 and so must have become established from seeds after red deer vacated the area about 1960. Two factors seem to have controlled the extent and rate of this succession.

First, there had to be a reliable seed source close to the potential woodland site. Even species as well adapted to wind dispersal as pine and birch deposit most of their seeds within a short distance of the parent tree. Booth (1984), for example, states that 75% of Scots pine seeds fall within 18 m of the parent tree. At Cairn Gorm, the exponential decline in sapling density with increasing distance from the forest edge (Figure 3) illustrates the importance of local seed availability. About 60% of the pine saplings were located within 500 m of the forest and 88% were within 1 km. Isolated plants were found at up to 4 km from a possible seed source, the seeds presumably having been blown across snowfields in late winter. However, seed shortage clearly prevailed beyond a few hundred metres from the forest edge.

Second, there had to be suitable gaps for seed germination and seedling establishment. In the northern corries of Cairn Gorm, sapling density was actually greatest at 650-700 m, well above the assumed tree line (Figure 4). Indeed, saplings were more abundant at 800-850 m than they were at any level up to 550-600 m. This apparent anomaly probably reflects differences in the availability of gaps for pine establishment (French et al., 1997b). Pine seedlings require open, unshaded sites with easy root access to well-drained mineral soil (Carlisle and Brown, 1968). Such conditions are uncommon on unburnt ground at low altitudes in the Cairngorms because a blanket of tall heather and other dwarf shrubs often blocks light and causes a deep layer of raw humus to accumulate at the soil surface. However, as altitude and exposure increase, heather becomes dwarfed, the mor humus layer becomes shallower, increasing areas of mineral soil are exposed and pine can establish.
Lack of suitable gaps can limit tree establishment unless they are continuously created. Pine began to colonise at Cairn Gorm in 1960, establishment rose to a peak around 1970 and then declined progressively to 1985 (French et al., 1997b). It seems that most available gaps had been filled by the 1980s and that future establishment would be much slower. An initial surge followed by a decline is quite typical of the pattern expected during colonisation by a single species. A return to the rapid rates of pine establishment of the 1960s would require continual and extensive disturbance of vegetation and soil to create new gaps.
Thus, given a seed source, conditions for pine establishment in the Cairngorms often may be more favourable at 550-750 m than they are on unburnt lower ground. Clearly a well-grown forest will not develop at such an altitude. However French et al. (1997a) have predicted the formation at Cairn Gorm of a sub-montane scrub similar to that already existing at Creag Fhiaclach within the next 100 years. Deliberate disturbance might accelerate the process.
Facilitating Woodland Regeneration
Clearly, where woodland regeneration is desired, the first priority in the Cairngorms is to drastically reduce red deer densities, at least to the levels indicated in Table 1. This could be achieved by fencing or by culling.
The practicality of fencing to encourage natural regeneration has been considered recently (e.g. Hester and Miller, 1995; Staines, 1995; Ratcliffe, 1998). There seems to be a consensus that fencing may be useful as a means of excluding deer from small blocks but that it is less desirable for preventing access to large areas. Fences are expensive both to erect and to maintain, particularly at remote upland sites, and capercaillie (Tetrao urogallus) and black grouse (T. tetrix) fatalities can occur (Catt et al, 1994). But they can also inhibit regeneration. The total exclusion of deer is likely to increase competition from the ungrazed vegetation and to accelerate the accumulation of surface litter. At the same time gaps for seedling establishment are no longer created by trampling and defecation (e.g. Halliday, 1998). Ungrazed grassy vegetation can encourage the proliferation of small mammals, including voles, which eat tree seeds, seedlings and small saplings (Miles and Kinnaird, 1979). Management for tree regeneration therefore should involve a degree of herbivore access to the site, perhaps on a seasonal basis.
After allowing red deer numbers to increase steadily for several decades, some landowners have been culling heavily in long-established saplings kept in check by repeated browsing recent years. Up until 1995, no appreciable reduction in deer numbers had been achieved within the Cairngorms/West Grampians area as a whole, where densities have been 13-14 per km2 since 1983 (Youngson and Stewart, 1996). At the local scale, however, culling has substantially reduced deer stocks at Inshriach, Creag Meagaidh and Abernethy to 5 per km2 or fewer, and this has allowed tree regeneration at these sites (Staines et al., 1995; Beaumont et al., 1995). At Caim Gorm, persistent human interference causing deer to abandon the area has also allowed regeneration to occur. However the extent, rate and longevity of any response can be limited by shortages both of seeds and of suitable gaps for seedling establishment.
Conclusion
In the Cairngorms, ground that has been left unburnt for several decades is likely to be covered by tall, dense heather and grass, with a thick carpet of bryophytes overlying a deep layer of raw humus. Such conditions are generally unsuitable for rapid seedling establishment by our small-seeded upland tree and shrub species. In these circumstances, forest development may start primarily with the growth of long-established saplings kept in check by repeated browsing (Figure 2). Reducing red deer densities by culling or by erecting fences may not in itself be enough to stimulate widespread and rapid regeneration. In the Cairngorms, consideration should also be given to such measures as controlled seasonal grazing, prescribed burning, scarifying and, in some circumstances, to the introduction of saplings or even seeds. Otherwise regeneration is likely to be slow, uncertain and localised.
References
Beaumont, D, Dugan, D, Evans, G and Taylor, S (1995): Deer management and tree regeneration in the RSPB Reserve at Abernethy Forest. Scottish Forestry 49, 155-161 .
Booth, T C (1984): Natural regeneration in the native pinewoods of Scotland. Scottish Forestry 38, 33-42.
Errant, J P, Clausen, T and Kuropat, P J (1985): Interactions of snowshoe hare and feltleaf willow in Alaska. Ecology 66, 1564-1573.
Bryant, J P, Kuropat, P J, Reichardt, P B and Clausen, T P (1991): Controls over the allocation of resources by woody plants to chemical antiherbivore defense. In Plant Defenses against Mammalion Herbivory (Ed. by R T Palo and C T Robbins), pp 84-102. CRC Press, Boca Raton, USA.
Cairngorms Working Party (1992): Common Sense and Sustainability: a partnership for the Cairngorms. HMSO, Edinburgh.
Canham, C D, McAninch, J B and Wood, D M (1994): Effects of frequency, timing and intensity of simulated browsing on growth and mortality of tree seedlings. Canadian journal of Forest Research 24, 817-825.
Carlisle, A (1977): The impact of man on the native pinewoods of Scotland. In Native Pinewoods of Scotland (Ed. by R G H Bunce and J N R Jeffers), pp 70-77. Institute of Terrestrial Ecology, Cambridge.
Carlisle, A and Brown, A H F (1968): Biological Flora of the British Isles: Pinus sylvestris L Journal of Ecology 56, 269-307.
Catt, D C, Duggan, D, Green, R E, Moncrieff, R, Moss, R, Picozzi, N, Summers, R W and Tyler, G A (1994): Collisions against fences by woodland grouse in Scotland. Forestry 67, 105-118.
Chapin, F S (1980): The mineral nutrition of wild plants. Annual Review of Ecology and Systematics 11, 233-260.
Cummins, R P and Miller, G R (1982): Damage by red deer (Cervus elaphus) enclosed in planted woodland. Scottish Forestry 36, 1-8.
Danell, K (1989): Elk browsing in pine stands: the biological background. Skogs och Londtbruksokodemiens Tidskrift 128, 105-109.
Danell, K, Niemala, P, Varvikko, T and Vuorisala, T (1991): Moose browsing on Scots pine along a gradient of plant productivity. Ecology 72, 1624-1633.
Darling, F F (1947): Natural History in the Highlands and Islands. Collins, London.
Edenius, L, Danell, K and Nyguist, H (1994): Effects of simulated moose browsing on growth, mortality and fecundity in Scots pine: relations to plant productivity. Canadian Journal of Forest Research 25, 529-535.
Fitter, A H and Jennings, R D (1975): The effects of sheep grazing on the growth and survival of seedling junipers (/uniperus communis L.). Journal of Applied Ecology 12,637-642.
Forrest, G I (1980): Genotypic variation among native Scots pine populations in Scotland based on monoterpene analysis. Forestry 53, 1 0 1 – 1 2 8 .
French, D D, Cummins, R P, Miller, G R, Bayfield, N G and McGowan, G M (I997a): Subalpine pine scrub in the Cairngorms. In The Ecology and Restoration of Montane and Subalpine Scrub Habitats in Scotland (Ed. by D Gilbert, D Horsfield and D B A Thompson), pp 81-89. Scottish Natural Heritage Review 83. SNH, Perth.
French, D D, Miller, G R and Cummins, R P (I997b): Recent development of high altitude Pinus sylvestris scrub in the northern Cairngorm Mountains. Biological Conservation 79, 133-144.
Gill, R M A (I992a): A review of damage by mammals in north temperate forests: 1. Deer. Forestry 65, 145-169.
Gill, R M A (I992b): A review of damage by mammals in north temperate forests: 3. Impact on trees and forests. Forestry 65, 363-388.
Halliday, J B (1998): Taynish NNR, woodland regeneration monitoring, bracken experiment. Native Pinewoods Discussion Group Newsletter 23, 26-27.
Heslop, R E (1981) : Soils. In The Cairngorms, 2nd edn. (Ed. by D Nethersole-Thompson and A Watson), pp 222-227. The Melven Press, Perth.
Hester, A J (1995): Scrub in the Scottish Uplands. Scottish Natural Heritage Review 24. SNH, Perth.
Hester, A J and Miller, G R (1995): Scrub and woodland regeneration: prospects for the future. In Heaths and Moorland: Cultural Landscapes (Ed. by DBA Thompson, A J Hester and M B Usher), pp 140-153. HMSO, London.
Hobbs, R J, Mallik, A U and Gimingham, C H (1984): Studies on fire in Scottish heathland communities. III. Vital attributes of the species. Journal of Ecology 72, 963-976.
Holloway, C W (1967): The effects of red deer and other animals on the natural regeneration of Scots pine. Unpublished PhD thesis, University of Aberdeen.
Kinnaird, J W (1974): Effect of site conditions on the regeneration of birch (Betufa pendula Roth and B. pubescens Ehrh.). Journal of Ecology 62, 467-472.
Laine, J and Mannerkoski, H (1980): Effect of fertilisation on tree growth and elk damage in young Scots pine stands planted on drained, nutrient-poor open bogs. Acta Forestalia Fennica 166, 1-45.
Macmillan, D C, Towers, S, MacLeay, S and Kupiec, J (1997): Modelling the potential distribution of the native woodland resource in the Cairngorms. Scottish Forestry 51, 70-75.
Mayle, B A and Staines, B W (1998): An overview of methods used for estimating the size of deer populations in Great Britain. In Population Ecology, Management and Welfare of Deer (Ed. by C R Goldspink, S King and R J Putman), pp 19-31. The Manchester Metropolitan University, Manchester.
Miles, J and Kinnaird, J W (1979): Grazing: with particular reference to birch, juniper and Scots pine in the Scottish Highlands. Scottish Forestry 33,280-288.
Miller, G R and Cummins, R P (1982): Regeneration of Scots pine Pinus sylvestris at a natural tree-line in the Cairngorm Mountains, Scotland. Holarctic Ecology 5, 27-34.
Miller, G R and Cummins, R P (1998): Browsing by red deer on naturally regenerated birch and juniper saplings on wintering ground at Glen Feshie. Scottish Forestry 52(3).
Miller, G R, Kinnaird, J W and Cummins, R P (1982): Liability of saplings to browsing on a red deer range in the Scottish Highlands. Journal of Applied Ecology 19, 941-951.
Mitchell, B and McCowan, D (1980): Estimating and comparing densities of red deer Cervus elaphus L. in concealing habitats. Institute of Terrestrial Ecology Annual Report for 1979, pp 7-13. Institute of Terrestrial Ecology, Cambridge.
Mitchell, B, McCowan, D and Willcox, N A (1982): Effects of deer in a woodland restoration enclosure. Scottish Forestry 36, 1 0 2 – 1 1 2 .
Mitchell, B, Staines, B W and Welch, D (1977): Ecology of Red Deer. Institute of Terrestrial Ecology, Cambridge.
Niemela, P and Danell, K (1988): Comparison of moose browsing on Scots pine (Pinus sylvestris) and lodgepole pine (P. contorta). Journal of Applied Ecology 25, 761-775.
Ouellet, J P, Boutin, S and Heard, S (1994): Responses to simulated grazing and browsing of vegetation available to caribou in the Arctic. Canadian Journal of Zoology 72, 1426-1435.
Palmer, S C F, Paterson, I S, Marquiss, M and Staines, B W (1998): The impact of deer browsing on regeneration of Scots pine: a preliminary study. In Population Ecology, Management and Welfare of Deer (Ed. by C R Goldspink, S King and R J Putman), pp 48-53. The Manchester Metropolitan University, Manchester.
Ratcliffe, P R (1998): Woodland deer management: integrating the control of their impact with multiple objective forest management in Scotland. In Population Ecology, Management and Welfare of Deer (Ed. by C R Goldspink, S King and R J Putman), pp 67-72. The Manchester Metropolitan University, Manchester.
Rusterholz, M and Turner, D C (1978): Experiments on nutritional wisdom of roe deer. Revue Suisse de Zoologie 85, 718-729.
Schwartz, C C, Nagy, J G and Regelin, W L (I980a): Juniper oil yield, terpenoid concentration, and antimicrobial effects on deer. Journal of Wildlife Management 44, 1 0 7 – 1 1 3 .
Schwartz, C C, Regelin, W L and Nagy, J G (1980b): Deer preference for juniper forage and volatile oil treated foods. Journal of Wildlife Management 44, 114-120.
Staines, B W (1995): The impact of red deer on the regeneration of native pinewoods. In Our Pinewood Heritage (Ed. by J R Aldhous), pp 107-114 . Forestry Commission, Farnham.
Staines, B W, Balharry, R and Welch, D (1995): The impact of red deer and their management on the natural heritage in the uplands. In Heaths and Moorlands: Cultural Landscapes (Ed. by D B A Thompson, A J Hester and M B Usher), pp 294-308. HMSO, London.
Stewart, F (1996): The effects of red deer (Cervus elaphus) on the regeneration of birch (Betula pubescens) woodland in the Scottish Highlands. Unpublished PhD thesis, University of Aberdeen.
Tahvanainen, J, Niemala, P and Henttonen, H (1991): Chemical aspects of herbivory in boreal forest-feeding by small rodents, hares and cervids. In Plant Defenses against Mammalian Herbivory (Ed. by R T Palo and C T Robbins), pp 115-131. CRC Press, Boca Raton, USA.
Whitmore, T C (1990): An Introduction to Tropical Rain Forests. Clarendon Press, Oxford.
Youngson, R W and Stewart, L K (1996): Trends in red deer populations within the Cairngorms core area. Botanical Journal of Scotland 48, 11l-l16.
